Vision is
by far the most important sensory modality
subserving spatial orientation, especially so in moving vehicles
such as aircraft. Without it, flight as we know it would be impossible,
whereas this would not necessarily be the case in the absence
of the vestibular or other sensory systems that provide orientation
information. For the most part, the function of vision in spatial
orientation is obvious, so a discussion proportional in size to
the importance of that function in orientation will not be presented
here. Certain special features of visual orientation deserve mention,
however. First, there are actually two separate visual systems,
and they have two distinct functions: object recognition and spatial
orientation. A knowledge of these systems is extremely
important,
both to help in understanding visual illusions in flight and to
appreciate the difficulties inherent in using flight instruments
for spatial orientation. Second, visual and vestibular orientation
information are integrated at very basic neural levels. For that
reason, spatial disorientation frequently is not amenable to correction
by higher-level neural processing.
Anatomy
of the Visual System
General
The retina,
an evaginated portion of the embryonic brain, consists of an outer
layer of pigmented epithelium and an inner layer of neural tissue.
Contained within the latter layer are the sensory rod and cone
cells, the bipolar and horizontal cells that comprise the intraretinal
afferent pathway from the rods and cones, and the multipolar ganglion
cells, the axons of which are the fibers of the optic nerve. The
cones, which number approximately 7 million in the human eye,
have a relatively high threshold to light energy. They are responsible
for sharp visual discrimination and color vision. The rods, of
which there are over 100 million, are much more sensitive to light
than the cones; they provide the ability to see in twilight and
at night. In the retinal macula, near the posterior pole of the
eye, the cone population achieves its greatest density; within
the macula, the fovea centralis--a small pit totally comprised
of tightly packed slender cones--provides the sharpest visual
acuity and is the anatomic basis for foveal, or central, vision.
The remainder of the eye is capable of far less visual acuity
and subserves paracentral and peripheral vision.
Having dendritic
connections with the rods and cones, the bipolar cells provide
axons that synapse with the dendrites or cell bodies of the multipolar
ganglion cells, whose axons in turn course parallel to the retinal
surface and converge at the optic disc. Emerging from the eye
as the optic nerve, they meet their counterparts from the opposite
eye in the optic chiasm and then continue in one of the optic
tracts, most likely to terminate in a lateral geniculate body,
but possibly in a superior colliculus or the pretectal area. Second
order neurons from the lateral geniculate body comprise the geniculocalcarine
tract, which becomes the optic radiation and terminates in the
primary visual cortex, the striate area of the occipital cerebral
cortex (Area 17). In the visual cortex, the retinal image is represented
as a more or less point-to-point projection from the lateral geniculate
body, which receives a similarly topographically structured projection
from both retinas. The lateral geniculate body and the primary
visual cortex are thus structurally and functionally suited for
the recognition and analysis of visual images. The superior colliculi
project to the visual association areas (Areas 18 and 19) of the
cerebral cortex via the pulvinar, and also eventually to the motor
nuclei of the extraocular muscles and muscles of the neck, and
appear to provide a pathway for certain gross ocular reflexes
of visual origin. Fibers entering the pretectal area are involved
in pupillary reflexes. In addition, most anatomic and physiologic
evidence indicates that information from the occipital visual
association areas, parietal cerebral cortex, and frontal eye movement
area (Area 8) is relayed through the paramedian pontine reticular
formation to the nuclei of the cranial nerves innervating the
extraocular muscles. Via this pathway and perhaps others involving
the superior colliculi, saccadic (fast) and pursuit (slow) eye
movements are initiated and controlled.
Visual-Vestibular
Convergence
Vision in
humans and other primates is highly dependent on cerebral cortical
structure and function, whereas vestibular orientation primarily
involves more primitive anatomic structures. Yet visual and vestibular
orientational processes are by no means independent. We know that
visually perceived motion information and probably other visual
orientational data reach the vestibular nuclei in the brain stem2,3,
but it appears that the integration of visual and vestibular information
is to a large extent accomplished in the cerebral cortex of humans.
The geniculostriate
projection system is divided both anatomically and functionally
into two parts: that incorporating the parvocellular layers of
the lateral geniculate body (the "parvo" system) and that incorporating
the magnocellular layers (the "magno" system). These systems are
largely segregated in the primary visual cortex, undergo further
segregation in the visual association cortex, and ultimately terminate
in the temporal and parietal lobes, respectively. The parvo system
neurons have smaller, more centrally located receptive fields
that exhibit high spatial resolution (acuity), and they respond
well to color. They do not, however, respond well to rapid
motion or high flicker rates. The magno cells, by comparison,
have larger receptive fields and respond better to motion and
flicker, but are relatively insensitive to color differences.
Magno neurons generally exhibit poorer spatial resolution, although
they seem to respond better than parvo neurons at low luminance
contrasts. In general, the parvo system is better at detecting
small, slowly moving, colored targets located near the center
of the visual field, while the magno system is more capable of
processing rapidly moving and optically degraded stimuli across
larger regions of the visual field.
What is
important about these two components of the geniculostriate system
is that the parvo system projects ventrally to the inferior temporal
areas, which are involved in visual search, pattern recognition,
and visual object memory, while the magno system projects dorsally
to the posterior parietal and superior temporal areas, which are
specialized for motion information processing. The cerebral cortical
areas to which the parvo system projects receive virtually no
vestibular afferents; the areas to which the magno system projects,
on the other hand, receive significant vestibular and other sensory
inputs, and are believed to be highly involved with maintaining
spatial orientation.
The posterior
parietal region projects heavily to cells of the pontine nuclei,
which in turn provide the mossy-fiber visual input to the cerebellar
cortex. Via the accessory optic and central tegmental tracts,
visual information also reaches the inferior olives, which provide
climbing fiber input to the cerebellar cortex. The cerebellar
cortex, specifically its flocculonodular lobe and vermis, also
receives direct mossy-fiber input from its vestibular system.
Thus, the cerebellum is another area of very strong visual-vestibular
convergence. Furthermore, the cerebellar Purkinje cells have inhibitory
connections in the vestibular nuclei and possibly even in the
vestibular end-organs; so visual-vestibular interactions mediated
by the cerebellum also occur at the level of the brain stem, and
maybe even peripherally.
Finally,
there is a confluence of visual and vestibular pathways in the
paramedian pontine reticular formation. Integration of visual
and vestibular information in the cerebellum and brain stem appears
to allow visual control of basic equilibratory reflexes of vestibular
origin. As might be expected, there also are afferent vestibular
influences on visual system nuclei; these influences have been
demonstrated in the lateral geniculate body and especially the
superior colliculus.
Visual
Information Processing
Primary
control of the human ability to move and orient oneself in three-dimensional
space is mediated by the visual system, as exemplified by the
fact that individuals without functioning vestibular systems ("labyrinthine
defectives") have virtually no problems with spatial orientation
unless they are deprived of vision. The underlying mechanisms
of visual orientation- information processing are revealed by
receptive field studies, which have been accomplished for the
peripheral retina, nuclear relays, and primary visual cortex.
Basically, these studies show that there are several types of
movement-detecting neurons and that these neurons respond differently
to the direction of movement, velocity of movement, size of the
stimulus, its orientation in space, and the level of illumination.
(For an excellent review of this fascinating topic, see Grǘsser
and Grǘsser-Cornehls4.)
As evidenced
by the division of the primate geniculostriate system into two
separate functional entities, however, vision must be considered
as two separate processes. Some researchers emphasize the role
of the ventral (parvo) system in object recognition (the "what"
system) and that of the dorsal (magno) system in spatial orientation
(the "where" system); others categorize the difference in terms
of form (occipito-temporal) versus motion (occipito-parietal)
processing. A recent theory suggests that the dorsal system is
primarily involved in processing information in peripersonal (near)
space during reaching and other visuomotor activity, whereas the
ventral system is principally engaged in visual scanning
in extrapersonal
(far) visual space.5
In the present discussion, we shall refer to the two systems as
the "focal" and "ambient" visual systems, respectively, subserving
the focal and ambient modes of visual processing. Certain aspects
of yet another visual process, the one responsible for generating
eye movements, will also be described.
Focal
Vision
Liebowitz
and Dichgans6 have
provided a very useful summary of the characteristics of
focal vision:
[The focal
visual mode] is concerned with object recognition and identification
and in general answers the question of "what." Focal vision involves
relatively fine detail (high spatial frequencies) and is correspondingly
best represented in the central visual fields. Information processed
by focal vision is ordinarily well represented in consciousness
and is critically related to physical parameters such as stimulus
energy and refractive error.
Focal vision
uses the central 30 degrees or so of the visual field. While it
is not primarily involved with orienting the individual in the
environment, it certainly contributes to conscious percepts of
orientation, such as those derived from judgments of distance
and depth and those obtained from reading flight instruments.
Tredici7
categorized the visual cues to distance and depth as monocular
or binocular. The monocular cues are (1) size constancy, the size
of the retinal image in relation to known and comparative sizes
of objects; (2) shape constancy, the shape of the retinal image
in relation to the known shape of the object (e.g., the foreshortening
of the image of a known circle into an ellipsoid shape means one
part of the circle is farther away than another); (3) motion parallax
(also called optical flow), the greater displacement of retinal
images of nearer objects when an individual is moving linearly
in the environment; (4) interposition, the partial obstruction
from view of more distant objects by nearer ones; (5) texture
or gradient, the apparent loss of detail with greater distance;
(6) linear perspective, the convergence of parallel lines at a
distance; (7) illumination perspective, which results from the
tendency to perceive the light source to be above an object and
from the association of more deeply shaded parts of an object
with being farther from the light source; and (8) aerial perspective,
the perception of objects to be more distant when the image is
relatively bluish or hazy.
The binocular
cues to depth and distance are (1) stereopsis, the visual appreciation
of three-dimensional space that results from the fusion of slightly
dissimilar retinal images of an object; (2) vergence, the medial
rotation of the eyes and the resulting direction of their gaze
along more or less converging lines, depending on whether the
viewed object is closer or farther, respectively; and (3) accommodation,
or focusing of the image by changing the curvature of the lens
of the eye. Of all the cues listed, size and shape constancy and
motion parallax appear to be most important for deriving distance
information in flying, and they are available at and well beyond
the distances at which binocular cues are useful. Stereopsis can
provide orientation information at distances up to only about
200 m; it is, however, more important in orientation than vergence
and accommodation, which are useless beyond about 6 m.
Ambient
Vision
Liebowitz
and Dichgans6 have also
provided a summary of ambient vision:
The ambient
visual mode subserves spatial localization and orientation and
is in general concerned with the question of "where." Ambient
vision is mediated by relatively large stimulus patterns so that
it typically involves stimulation of the peripheral visual field
and relatively coarse detail (low spatial frequencies). Unlike
focal vision, ambient vision is not systematically related to
either stimulus energy or optical image quality. Rather, provided
the stimulus is visible, orientation responses appear to be elicited
on an "all or none" basis The conscious concomitant of ambient
stimulation is low or frequently completely absent.
Ambient
vision, therefore, is primarily involved with orienting the individual
in the environment. Furthermore, this function is largely independent
of the function of focal vision. This becomes evident in view
of the fact that one can fully occupy central vision with the
task of reading while simultaneously obtaining sufficient orientation
cues with peripheral vision to walk or ride a bicycle. It is also
evidenced by the ability of certain patients with cerebral cortical
lesions to maintain visual orientation responses even though their
ability to discriminate objects is lost.
While we
commonly think of ambient vision as dependent on stimulation of
peripheral visual fields, it is more accurate to consider ambient
vision as involving large areas of the total visual field, which
of course must include the visual periphery. In other words, ambient
vision is not so much location- dependent as it is area-dependent.
Moreover, ambient vision is stimulated much more effectively by
large images or groups of images perceived to be at a distance
than by those appearing to be close.
The function
of ambient vision in orientation can be thought of as two processes,
one providing motion cues and the other providing position cues.
Large, coherently moving contrasts detected over a large area
of the visual field result in vection, i.e., a visually induced
percept of self-motion. If the moving contrasts revolve relative
to the subject, he or she perceives rotational self-motion or
angular vection (also called circular vection), which can be in
the pitch, roll, yaw, or any intermediate plane. If the moving
contrasts enlarge and diverge from a distant point, become smaller
and converge in the distance, or otherwise indicate linear motion,
the percept of self-motion that results is linear vection, which
also can be in any direction. Vection can, of course, be veridical
or illusory, depending on whether actual or merely apparent motion
of the subject is occurring. One can appreciate the importance
of ambient vision in orientation by recalling the powerful sensations
of self-motion generated by certain scenes in wide-screen motion
pictures (e.g., flying through the Grand Canyon in an IMAX theater).
Position
cues provided by ambient vision are readily evidenced in the stabilization
of posture that vision affords patients with defective vestibular
or spinal proprioceptive systems. The essential visual parameter
contributing to postural stability appears to be the motion of
the retinal image that results from minor deviations from one's
desired postural position. Visual effects on posture also can
be seen in the phenomenon of height vertigo. As the distance from
(height above) a stable visual environment increases, the amount
of body sway necessary for the retinal image movement to be above
threshold increases. Above a certain height, the ability of this
visual mechanism to contribute to postural stability is exceeded,
and vision indicates posture to be stable despite large body sways.
The conflict between visual orientation information, indicating
relative stability, and the vestibular and somatosensory data,
indicating large body sways, results in the unsettling experience
of vertigo.
One more
distinction between focal and ambient visual function should be
emphasized. In general, focal vision serves to orient the perceived
object relative to the individual, whereas ambient vision serves
to orient the individual relative to the perceived environment.
When both focal and ambient vision are present, orienting a focally
perceived object relative to the ambient visual environment is
easy, whether the mechanism employed involves first orienting
the object to oneself and then orienting oneself and the object
to the environment or involves orienting the object directly to
the environment. When only focal vision is available, however,
it can be difficult to orient oneself correctly to a focally perceived
environmental orientation cue because the natural tendency is
to perceive oneself as stable and upright and to perceive the
focally viewed object as oriented with respect to the stable and
upright egocentric reference frame. This phenomenon can cause
a pilot to misjudge his or her approach to a night landing, for
example, when only the runway lights and a few other focal visual
cues are available for spatial orientation.
Eye Movements
We distinguish
between two fundamental types of eye movement: smooth movements,
including pursuit, vergence, and those driven by the vestibular
system; and saccadic (jerky) movements. Smooth eye movements are
controlled at least in part by the posterior parietal cerebral
cortex and surrounding areas, as evidenced by functional deficits
resulting from damage to these areas. Eye movements of vestibular
origin are primarily generated by very basic reflexes involving
brain stem mechanisms; and because visual pursuit eye movements
are impaired by vestibular and certain cerebellar lesions, the
vestibular system appears to be involved in control of smooth
eye movements of visual origin. Saccadic eye movements are controlled
mainly by the frontal eye fields of the cerebral cortex, which
work with the superior colliculus in generating these movements.
The frontal eye fields receive their visual input from the cortical
visual association areas.
The maintenance
of visual orientation in a dynamic motional environment is greatly
enhanced by the ability to move the eyes, primarily because the
retinal image of the environment can be stabilized by appropriate
eye movements. Very powerful and important mechanisms involved
in reflexive vestibular stabilization of the retinal image will
be discussed in the section dealing with vestibular function.
Visual pursuit movements also serve to stabilize the retinal image,
as long as the relative motion between the head and the visual
environment (or object being observed in it) is less than about
600/sec: targets moving at higher relative velocities necessitate
either saccadic eye movements or voluntary head movements for
adequate tracking. Saccadic eye movements are used voluntarily
or reflexively to acquire a target, i.e., to move it into focal
vision, or to catch up to a target that cannot be maintained on
the fovea by pursuit movements. Under some circumstances, pursuit
and saccadic eye movements alternate in a pattern of reflexive
slow tracking and fast-back tracking called optokinetic nystagmus.
This type of eye-movement response is typically elicited in the
laboratory by surrounding the subject with a rotating striped
drum; however, one can exhibit and experience optokinetic nystagmus
quite readily in a more natural setting by watching railroad cars
go by while waiting at a railroad crossing. Movement of the visual
environment sufficient to elicit optokinetic nystagmus provides
a stimulus that can either enhance or compete with the vestibular
elicitation of eye movements, depending on whether the visually
perceived motion is compatible or incompatible, respectively,
with the motion sensed by the vestibular system.
Vergence
movements, which aid binocular distance and motion perception
at very close range, are of relatively minor importance in spatial
orientation when compared with the image-stabilizing pursuit and
saccadic eye movements. Vergence assumes some degree of importance,
however, under conditions where a large visual environment is
being simulated in a confined space. Failure to account for vergence
effects can result in loss of simulation fidelity: a subject whose
eyes are converged to fuse an image representing a large, distant
object will perceive that object as small and near. To overcome
this problem, visual flight simulators display distant scenes
at the outer limit of vergence effects (7-10 meters) or use lenses
or mirrors to put the displayed scene at optical infinity.
Even though
gross stabilization of the retinal image aids object
recognition
and spatial orientation by enhancing visual acuity, absolute stability
of an image is associated with a marked decrease in visual acuity
and form perception8.
This stability-induced decrement is avoided by continual voluntary
and involuntary movements of the eyes, even during fixation of
an object. We are unaware of these small eye movements, however,
and the visual world appears stable.
Voluntary
scanning and tracking movements of the eyes are associated with
the appearance of a stable visual environment, but why this is
so is not readily apparent. Early investigators postulated that
proprioceptive information from the extraocular muscles provides
not only feedback signals for the control of eye movements but
also the afferent information needed to correlate eye movements
with retinal image movements and arrive at a subjective determination
of a stable visual environment. An alternative mechanism for oculomotor
control and the subjective appreciation of visual stability is
the "corollary discharge" or feed-forward mechanism first proposed
by Helmholtz and subsequently by Sperry9 and
others. Sperry concluded: "Thus, an excitation pattern that
normally results in a movement that will cause a displacement
of the visual image on the retina may have a corollary discharge
into the visual centers to compensate for the retinal displacement.
This implies an anticipatory adjustment in the visual centers
specific for each movement with regard to its direction and speed."
The theoretical aspects of visual perception of movement and stability
have been expanded over the years into various models based on
"inflow" (afference), "outflow" (efference), and even hybrid sensory
mechanisms. The interested reader will enjoy Cohen's concise discussion
of these models as they relate to spatial orientation10.
In developing
the important points on visual orientation, we have emphasized
the "focal-ambient" dichotomy. As visual science matures further,
this simplistic construct will likely be replaced by more complex
but valid models of visual processes. Presently we are enthusiastic
about a theory in which the dichotomy emphasized is that between
the peripersonal (near) and focal extrapersonal (far) visual realms.
This theory argues that the dorsal cortical system and its magno
projection pathways are more involved in processing visual information
from peripersonal space, while the ventral system and its parvo
projections attend to the focal extrapersonal visual environment.
The theory also suggests that visual attention is organized to
be employed more efficiently in some sectors of three-dimensional
visual space than in others (e.g., far vision is biased toward
the upper visual field and utilizes local form processing, while
near vision is biased toward the lower visual field and is better
at global form processing), and that ambient extrapersonal information
is largely excluded from attentional mechanisms. Certainly, the
current state of knowledge concerning visual orientation is fluid.
VESTIBULAR FUNCTION
The role of vestibular function
in spatial orientation is not so overt as that of vision but is
extremely important for three major reasons. First, the vestibular
system provides the structural and functional substrate for reflexes
that serve to stabilize vision when motion of the head and body
would otherwise result in blurring of the retinal image. Second,
the vestibular system provides orientational information with
reference to which both skilled and reflexive motor activities
are automatically executed. Third, the vestibular system provides,
in the absence of vision, a reasonably accurate percept of motion
and position, as long as the pattern of stimulation remains within
certain naturally occurring bounds. Because the details of vestibular
anatomy and physiology are not usually well known by medical professionals,
and because a working knowledge of them is essential to the understanding
of spatial disorientation in flight, these details will be presented
in the following sections.
Vestibular Anatomy
End-Organs
The vestibular end-organs are
smaller than most people realize, measuring just 1.5 cm across.
They reside well-protected within some of the densest bone in
the body, the petrous portion of the temporal bone. Each temporal
bone contains a tortuous excavation known as the bony labyrinth,
which is filled with perilymph, a fluid much like cerebrospinal
fluid in composition. The bony labyrinth consists of three main
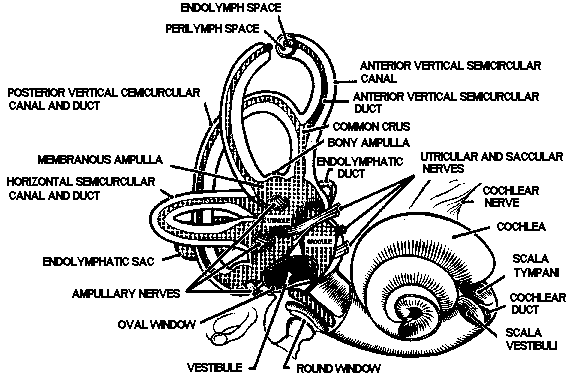
parts: the cochlea, the vestibule, and the semicircular canals
(Fig. 3). Within each part of the bony labyrinth is a part of
the delicate, tubular, membranous labyrinth, which contains endolymph,
a fluid characterized by its relatively high concentration of
potassium. In the cochlea, the membranous labyrinth is
called
the cochlear duct or scala media; this organ converts acoustic
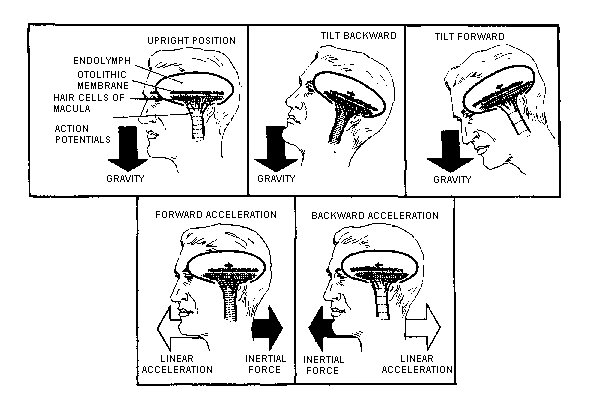
energy into neural information. In the vestibule lie the two otolith
organs, the utricle and the saccule. They translate gravitational
and inertial forces into spatial orientation information--specifically,
information about angular position (tilt) and linear motion of
the head. The semicircular ducts, contained in the semicircular
canals, convert inertial torques into information about angular
motion of the head. The three semicircular canals and their included
semicircular ducts are oriented in three mutually perpendicular
planes, thus inspiring the names of the canals: anterior vertical
(or superior), posterior vertical (or posterior), and horizontal
(or lateral).
The semicircular ducts communicate
at both ends with the utricle, and one end of each duct is dilated
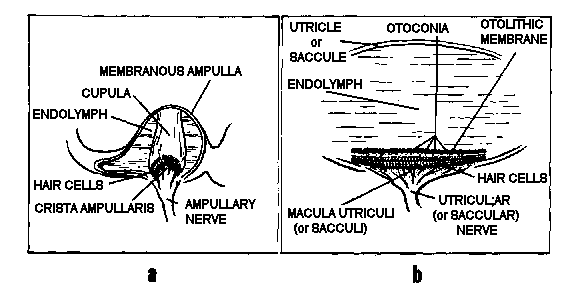
to form an ampulla. Inside each ampulla lies a crest of neuroepithelium,
the crista ampullaris. Atop the crista, occluding the duct, is
a gelatinous structure called the cupula (Fig. 4a). The hair cells
of which the crista ampullaris is composed project their cilia
into the base of the cupula, so that whenever inertial torques
of the endolymph ring in the semicircular duct deviate the cupula,
the cilia are bent.
Lining the bottom of the utricle
is another patch of neuroepithelium, the macula utriculi, whose
plane is close to horizontal except for a 20-30° upward slope
of its anterior end; and on the medial wall of the saccule in
an approximately vertical plane is still another, the macula sacculi
(Fig. 4b). The cilia of the hair cells comprising these structures
project into overlying otolithic membranes, one above each macula.
The otolithic membranes are gelatinous structures containing many
tiny calcium carbonate crystals, called otoconia, which are held
together by a network of connective tissue. Having almost three
times the density of the surrounding endolymph, the otolithic
membranes displace endolymph and shift position relative to their
respective maculae when subjected to changing gravitoinertial
forces. This shifting of the otolithic membrane position results
in bending of the cilia of the macular hair cells.
Figure
3. Gross anatomy of the inner ear. The bony semicircular canals
and vestibule contain the membranous semicircular ducts and otolith
organs, respectively.
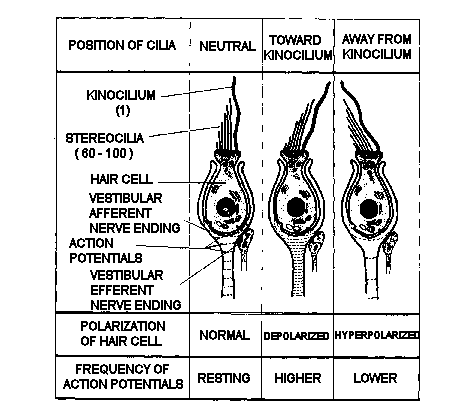
The hair cell is the functional
unit of the vestibular sensory system. It converts spatial and
temporal patterns of mechanical energy applied to the head into
neural information. Each hair cell possesses one relatively large
kinocilium on one side of the top of the cell and up to 100 smaller
stereocilia on the same surface. Hair cells thus exhibit morphologic
polarization, that is, they are oriented in a particular direction.
The functional correlate of this polarization is that when the
cilia of a hair cell are bent in the direction of its kinocilium,
the cell undergoes an electrical depolarization, and the frequency
of action potentials generated in the vestibular neuron attached
to the hair cell increases above a certain resting frequency;
the greater the deviation of the cilia, the higher the frequency.
Similarly, when its cilia are bent away from the side with the
kinocilium, the hair cell undergoes an electrical hyperpolarization,
and the frequency of action potentials in the corresponding neuron
in the vestibular nerve decreases (Fig. 5).
Figure
4. Vestibular end-organs. a. The ampulla of a semicircular
duct, containing the crista ampullaris and cupula. b. A representative
otolith organ, with its macula and otolithic membrane.
The same basic process just described
occurs in all the hair cells in the three cristae and both maculae;
the important differences lie in the physical events that cause
the deviation of cilia in the directions in which the various
groups of hair cells are oriented. The hair cells of a crista
ampullaris respond to the inertial torque of the ring of endolymph
contained in the attached semicircular duct as the reacting endolymph
exerts pressure on the cupula and deviates it. The hair cells
of a macula, on the other hand, respond to the gravitoinertial
force acting to displace the overlying otolithic membrane. As
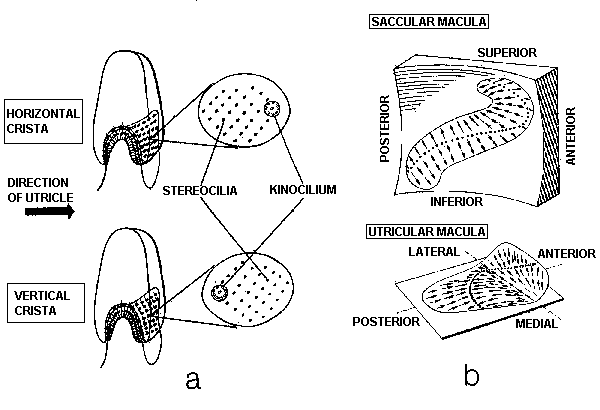
indicated in Figure 6a, all of the hair cells in the crista of
the horizontal semicircular duct are oriented so that their kinocilia
are on the side of the cell facing the utricle. Thus, utriculopetal
endolymphatic pressure on the cupula deviates the cilia of these
hair cells toward the kinocilia, and all the hair cells in the
crista depolarize. The hair cells in the cristae of the vertical
semicircular ducts are oriented in the opposite fashion; that
is, their kinocilia are all on the side away from the utricle.
In the ampullae of the vertical semicircular ducts, therefore,
utriculopetal endolymphatic pressure deviates the cilia away from
the kinocilia, causing all the hair cells in these cristae to
hyperpolarize. In contrast, the hair cells of the maculae are
not oriented unidirectionally across the neuroepithelium: the
direction of their morphologic polarization depends on where they
lie on the macula (Fig. 6b). In both maculae there is a central
line of reflection, on opposing sides of which the hair cells
assume an opposite orientation. In the utricular macula, the kinocilia
of the hair cells are all oriented toward this line of reflection;
whereas in the saccular macula, they are oriented away from it.
Because the line of reflection on each macula curves at least
90°, the hair cells, having morphologic polarization roughly perpendicular
to this line, exhibit virtually all possible orientations on the
plane of the macula. Thus, the orthogonality of the planes of
the three semicircular ducts enables them efficiently to detect
angular motion in any plane; and the perpendicularity of the planes
of the maculae plus the omnidirectionality of the orientation
of the hair cells in the maculae allow the efficient detection
of gravitoinertial forces acting in any direction.
Figure
5. Function of a vestibular hair cell. When mechanical forces
deviate the cilia toward the side of the cell with the kinocilium,
the hair cell depolarizes and the frequency of action potentials
in the associated afferent vestibular neuron increases. When the
cilia are deviated in the opposite direction, the hair cell hyperpolarizes
and the frequency of action potentials decreases.
Neural Pathways
To help the reader better organize
the potentially confusing vestibular neuroanatomy, a somewhat
simplified overview of the major neural connections of the vestibular
system is presented in Figure 7. The utricular nerve, two saccular
nerves, and the three ampullary nerves converge to form the vestibular
nerve, a portion of the VIIIth cranial or statoacoustic nerve.
Within the vestibular nerve lies the vestibular (or Scarpa's)
ganglion, which is composed of the cell bodies of the vestibular
neurons. The dendrites of these bipolar neurons invest the hair
cells of the cristae and maculae; most of their axons terminate
in the four vestibular nuclei in the brain stem--the superior,
medial, lateral, and inferior nuclei--but some axons enter the
phylogenetically ancient parts of the cerebellum to terminate
in the fastigial nuclei and in the cortex of the flocculonodular
lobe and other parts of the posterior vermis.
Figure
6. Morphologic polarization in vestibular neuroepithelia. a. All the hair
cells in the cristae of the horizontal semicircular
ducts are oriented so that their kinocilia are in the direction
of the utricle; those hair cells in the cristae of the vertical
ducts have their kinocilia directed away from the utricle. b.
The maculae of the saccule (above) and utricle (below) also exhibit
polarization: the arrows indicate the direction of the kinocilia
of the hair cells in the various regions of the maculae. (Adapted
from Spoendlin11. )
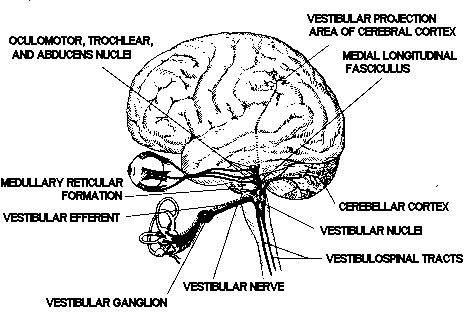
The vestibular nuclei project
via secondary vestibular tracts to motor nuclei of cranial and
spinal nerves and to the cerebellum. Because vestibulo-ocular
reflexes are a major function of the vestibular system, it is
not surprising to find ample projections from the vestibular nuclei
to the nuclei of the oculomotor trochlear, and abducens nerves
(cranial nerves III, IV, and VI, respectively). The major pathway
of these projections is the ascending medial longitudinal fasciculus
(MLF). The basic vestibulo-ocular reflex is thus served by sensor
and effector cells and an intercalated three-neuron reflex arc
from the vestibular nerve to the vestibular nuclei to the nuclei
innervating the extraocular muscles. In addition, indirect multisynaptic
pathways course from the vestibular nuclei through the paramedian
pontine reticular formation to the oculomotor and other nuclei.
The principle of ipsilateral facilitation and contralateral inhibition
via an interneuron clearly operates in vestibulo-ocular reflexes,
and numerous crossed internuclear connections provide evidence
of this. The vestibulo-ocular reflexes that the various ascending
and crossed pathways support serve to stabilize the retinal image
by moving the eyes in the direction opposite that of the motion
of the head.
Figure
7. Major connections and projections of the vestibular system.
Via the descending MLF and medial
vestibulospinal tract, crossed and uncrossed projections from
the vestibular nuclei reach the nuclei of the spinal accessory
nerve (cranial nerve XI) and motor nuclei in the cervical cord.
These projections form the anatomic substrate for vestibulocollic
reflexes, which serve to stabilize the head by appropriate action
of the sternocleidomastoid and other neck muscles. A third projection
is that from primarily the lateral vestibular nucleus into the
ventral gray matter throughout the length of the spinal cord.
This important pathway is the uncrossed lateral vestibulospinal
tract, which enables the vestibulospinal (postural) reflexes to
help stabilize the body with respect to an inertial frame of reference
by means of sustained and transient vestibular influences on basic
spinal reflexes.
Secondary vestibulocerebellar
fibers course from the vestibular nuclei into the ipsilateral
and contralateral fastigial nuclei and to the cerebellar cortex
of the flocculonodular lobe and elsewhere. Returning from the
fastigial and other cerebellar nuclei, crossed and uncrossed fibers
of the cerebellobulbar tract terminate in the vestibular nuclei
and in the associated reticular formation. There are also efferent
fibers from the cerebellum, probably arising in the cerebellar
cortex, that terminate not in nuclear structures but on dendritic
endings of primary vestibular afferent neurons in the vestibular
neuroepithelia. Such fibers are those of the vestibular efferent
system, which appears to modulate or control the information arising
from the vestibular end-organs. The primary and secondary vestibulocerebellar
fibers and those returning from the cerebellum to the vestibular
area of the brain stem comprise the juxtarestiform body of the
inferior cerebellar peduncle. This structure, along with the vestibular
end-organs, nuclei, and projection areas in the cerebellum, collectively
constitute the so-called vestibulocerebellar axis, the neural
complex responsible for processing primary spatial orientation
information and initiating adaptive and protective behavior based
on that information.
Several additional projections,
more obvious functionally than anatomically, are those to certain
autonomic nuclei of the brainstem and to the cerebral cortex.
The dorsal motor nucleus of cranial nerve X (vagus) and other
autonomic cell groups in the medulla and pons receive secondary
vestibular fibers, largely from the medial vestibular nucleus;
these fibers mediate vestibulovegetative reflexes, which are manifested
in the symptoms of motion sickness (pallor, perspiration, nausea,
and vomiting) that can result from excessive or otherwise abnormal
vestibular stimulation. Via vestibulothalamic and thalamocortical
pathways, vestibular information eventually reaches the primary
vestibular projection area of the cerebral cortex, located
in
the parietal and parieto-temporal cortex. This projection area
is provided with vestibular, visual, and somatosensory (proprioceptive)
inputs and is evidently associated with spatial orientation processing
and with integration of higher-order sensorimotor activity. In
addition, vestibular information can be transmitted via long polysynaptic
pathways through the brain stem reticular formation and medial
thalamus to wide areas of the cerebral cortex; the nonspecific
cortical responses to vestibular stimuli that are evoked via this
pathway appear to be associated with an arousal or alerting mechanism.
Vestibular Information Processing
As the reader probably deduced
while reading the discussion of the anatomy of the vestibular
end-organs, angular accelerations are the adequate (that is, physiologic)
stimuli for the semicircular ducts, and linear accelerations and
gravity are the adequate stimuli for the otolith organs. This
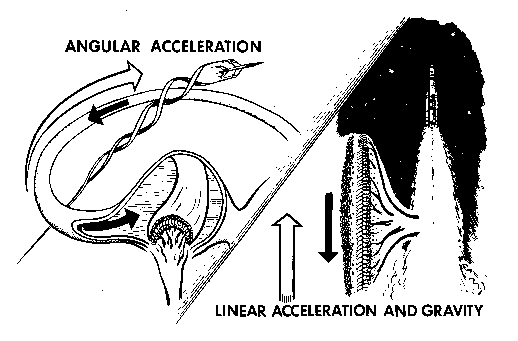
statement, illustrated in Figure 8, is the cardinal principle
of vestibular mechanics. How the reactive torques and gravitoinertial
forces stimulate the hair cells of the cristae and maculae, respectively,
and produce changes in the frequency of action potentials in the
associated vestibular neurons has already been discussed. The
resulting frequency-coded messages are transmitted into the several
central vestibular projection areas as raw orientational data
to be further processed as necessary for the various functions
served by such data. These functions are the vestibular reflexes,
voluntary movement, and the perception of orientation.
Figure
8. The cardinal principle of vestibular mechanics: angular accelerations
stimulate the semicircular ducts; linear accelerations and gravity
stimulate the otolith organs.
Vestibular Reflexes
As stated so well by G. Melvill
Jones12, "...for control of eye movement relative to
space the motor outflow can operate on three fairly discrete anatomical
platforms, namely: (1) the eye-in-skull platform, driven by the
external eye muscles rotating the eyeball relative to the skull;
(2) the skull-on-body platform driven by the neck muscles; and
(3) the body platform, operated by the complex neuromuscular mechanisms
responsible for postural control."
In humans, the retinal image is
stabilized mainly by vestibulo-ocular reflexes, primarily those
of semicircular-duct origin. A simple demonstration can help one
appreciate the contribution of the vestibulo-ocular reflexes to
retinal-image stabilization. Holding the extended fingers half
a meter or so in front of the face, one can move the fingers slowly
from side to side and still see them clearly because of visual
(optokinetic) tracking reflexes. As the rate, or correspondingly,
the frequency, of movement becomes greater, one eventually reaches
a point where the fingers cannot be seen clearly--they are blurred
by the movement. This point is at about 60°/sec or 1 to 2 Hz for
most people. Now, if the fingers are held still and the head is
rotated back and forth at the frequency at which the fingers became
blurred when they were moved, the fingers remain perfectly clear.
Even at considerably higher frequencies of head movement, the
vestibulo-ocular reflexes initiated by the resulting
stimulation
of the semicircular ducts function to keep the image of the fingers
clear. Thus, at lower frequencies of movement of the external
world relative to the body or vice versa, the visual system stabilizes
the retinal image by means of optokinetic reflexes. As the frequencies
of such relative movement become greater, however, the vestibular
system, by means of vestibulo-ocular reflexes, assumes progressively
more of this function; and at the higher frequencies of relative
motion characteristically generated only by motions of the head
and body, the vestibular system is responsible for stabilizing
the retinal image.
The mechanism by which stimulation
of the semicircular ducts results in retinal image stabilization
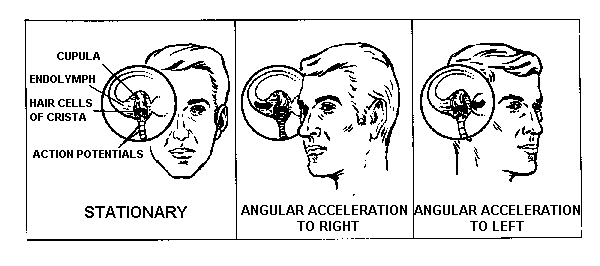
is simple, at least conceptually (Fig. 9). When the head is turned
to the right in the horizontal (yaw) plane, the angular acceleration
of the head creates a reactive torque in the ring of endolymph
in (mainly) the horizontal semicircular duct. The reacting endolymph
then exerts pressure on the cupula, deviating the cupula in the
right ear in a utriculopetal direction, depolarizing the hair
cells of the associated crista ampullaris and increasing the frequency
of the action potentials in the corresponding ampullary nerve.
In the left ear, the endolymph deviates the cupula in a utriculofugal
direction, thereby hyperpolarizing the hair cells and decreasing
the frequency of the action potentials generated. As excitatory
neural signals are relayed to the contralateral lateral rectus
and ipsilateral medial rectus muscles, and inhibitory signals
are simultaneously relayed to the antagonists, a conjugate deviation
of the eyes results from the described changes in ampullary neural
activity. The direction of this conjugate eye deviation is thus
the same as that of the angular reaction of the endolymph, and
the angular velocity of the eye deviation is proportional to the
pressure exerted by the endolymph on the cupula. The resulting
eye movement is, therefore, compensatory; that is, it adjusts
the angular position of the eye to compensate for changes in angular
position of the head and thereby prevents slippage of the retinal
image over the retina. Because the amount of angular deviation
of the eye is physically limited, rapid movements of the eye in
the direction opposite the compensatory motion are employed to
return the eye to its initial position or to advance it to a position
from which it can sustain a compensatory sweep for a suitable
length of time. These rapid eye movements are anticompensatory,
and because of their very high angular velocity, motion is not
perceived during this phase of the vestibulo-ocular reflex.
Figure
9. Mechanism of action of a horizontal semicircular duct and the
resulting reflex eye movement. Angular acceleration to the
right increases the frequency of action potentials originating
in the right ampullary nerve and decreases those in the left.
This pattern of neural signals causes extraocular muscles to rotate
the eyes in the direction opposite that of head rotation, thus
stabilizing the retinal image with a compensatory eye movement.
Angular acceleration to the left has the opposite effect.
With the
usual rapid, high-frequency rotations of the head, the rotational
inertia of the endolymph acts to deviate the cupula as the angular
velocity of the head builds, and the angular momentum gained by
the endolymph during the brief acceleration acts to drive the
cupula back to its resting position when the head decelerates
to a stop. The cupula-endolymph system thus functions as an integrating
angular accelerometer, that is, it converts angular, acceleration
data into a neural signal proportional to the angular velocity
of the head. This is true for angular accelerations occurring
at frequencies normally encountered in terrestrial activities;
when angular accelerations outside the dynamic response range
of the cupula-endolymph system are experienced, the system no
longer provides accurate angular velocity information.
When angular
accelerations are relatively sustained or when a cupula is kept
in a deviated position by other means, such as caloric testing,
the compensatory and anticompensatory phases of the vestibulo-ocular
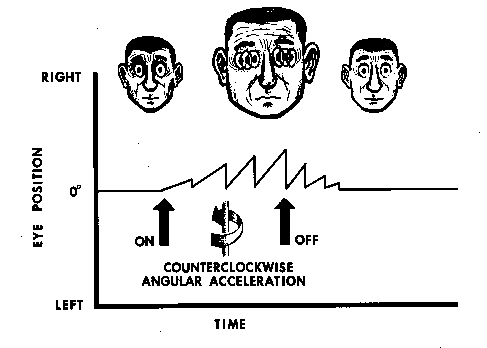
reflex are repeated, resulting in beats of ocular nystagmus (Fig.
10). The compensatory phase of the vestibulo-ocular reflex is
then called the slow phase of nystagmus, and the anticompensatory
phase is called the fast or quick phase. The direction of the
quick phase is used to label the direction of the nystagmus because
the direction of the rapid motion of the eye is easier to detect
clinically. The vertical semicircular ducts operate in an analogous
manner, with the vestibulo-ocular reflexes elicited by their stimulation
being appropriate to the plane of the angular acceleration resulting
in that stimulation. Thus, a vestibulo-ocular reflex with downward
compensatory and upward anti-compensatory phases results from
the stimulation of the vertical semicircular ducts by pitch-up
(-ay) angular acceleration, and with sufficient stimulation in
this plane, up-beating vertical nystagmus results. Angular accelerations
in the roll plane result in vestibulo-ocular reflexes with clockwise
and counterclockwise compensatory and anticompensatory phases
and in rotary nystagmus. Other planes of stimulation are associated
with other directions of eye movement such as oblique or horizonto-rotary.
Figure
10. Ocular nystagmus--repeating compensatory and anticompensatory
eye movements--resulting from vestibular stimulation. In this
case, the stimulation is a yawing angular acceleration to the
left, and the anticompensatory, or quick-phase, nystagmic response
is also to the left.
As should be expected, there also
are vestibulo-ocular reflexes of otolith-organ origin. Initiating
these reflexes are the shearing actions that bend the cilia of
macular hair cells as inertial forces or gravity cause the otolithic
membranes to slide to various positions over their maculae (Fig.
11). Each position that can be assumed by an otolithic membrane
relative to its macula evokes a particuiar spatial pattern of
frequencies of action potentials in the corresponding utricular
or saccular nerve, and that pattern is associated with a particular
set of compatible stimulus conditions such as backward tilt of
the head or forward linear acceleration. These patterns of action
potentials from the various otolith organs are correlated and
integrated in the vestibular nuclei and cerebellum with orientational
information from the semicircular ducts and other sensory modalities;
appropriate orientational percepts and motor activities eventually
result. Lateral (ay) linear accelerations can elicit horizontal
reflexive eye movements, including nystagmus, presumably as a
result of utricular stimulation. Similarly, vertical (az) linear
accelerations can elicit vertical eye movements, most likely as
a result of stimulation of the saccule; the term elevator reflex
is sometimes used to describe this response because it is readily
provoked by the vertical linear accelerations associated with
riding in an elevator. The utility of these horizontal and vertical
vestibulo-ocular reflexes of otolith-organ origin is readily apparent:
like the reflexes of semicircular- duct origin, they help stabilize
the retinal image. Less obvious is the usefulness of the ocular
countertorsion reflex (Fig. 12), which repositions the eyes about
their visual (anteroposterior) axes in response to the otolith-organ
stimulation resulting from tilting the head laterally in the opposite
direction. Presumably, this reflex contributes to retinal image
stabilization by providing a response to changing directions of
the force of gravity.
Our understanding of the vestibulocollic
reflexes has not developed to the same degree as our understanding
of the vestibulo-ocular reflexes, although some clinical use has
been made of measurements of rotation of the head on the neck
in response to vestibular stimulation. Perhaps this situation
reflects the fact that vestibulocollic reflexes are not as effective
as the vestibulo-ocular reflexes in stabilizing the retinal image,
at least not in humans. Such is not the case in other species,
however; birds exhibit extremely effective reflex control of head
position under conditions of bodily motion--even nystagmic head
movements are quite easy to elicit. The high level of development
of the vestibulocollic reflexes in birds is certainly either a
cause or a consequence of the relative immobility of birds' eyes
in their heads. Nonetheless, the ability of a human (or any other
vertebrate with a mobile head) to keep the head upright with respect
to the direction of applied gravitoinertial force is maintained
through tonic vestibular influences on the muscles of the neck.
Vestibulospinal reflexes operate
to ensure stability of the body. Transient linear and angular
accelerations, such as those experienced in tripping and falling,
provoke rapid activation of various groups of extensor and flexor
muscles to return the body to the stable position or at least
to minimize the ultimate effect of the instability. Everyone has
experienced the reflex arm movements that serve to break a fall,
and most have observed the more highly developed righting reflexes
that cats exhibit when dropped from an upside-down position; these
are examples of vestibulospinal reflexes. Less spectacular, but
nevertheless extremely important, are the sustained vestibular
influences on posture that are exerted through tonic activation
of so-called "antigravity" muscles such as hip and knee extensors.
These vestibular reflexes, of course, help keep the body upright
with respect to the direction of the force of gravity.
Figure
11. Mechanism of action of an otolith organ. A change in direction
of the force of gravity (above) or a linear acceleration (below)
causes the otolithic membrane to shift its position with respect
to its macula, thereby generating a new pattern of action potentials
in the utricular or saccular nerve. Shifting of the otolithic
membranes can elicit compensatory vestibulo-ocular reflexes and
nystagmus, as well as perceptual effects.
Voluntary Movement
It has been shown how the various
reflexes of vestibular origin serve to stabilize the body in general
and the retinal image in particular. The vestibular system is
also important in that it provides data for the proper execution
of voluntary movement. To realize just how important such vestibular
data are in this context, one must first recognize the fact that
skilled voluntary movements are preprogrammed; that is, once initiated,
they are executed according to a predetermined pattern and sequence,
without the benefit of simultaneous sensory feedback to the higher
neural levels from which they originate. The simple act of writing
one's signature, for example, involves such rapid changes in speed
and direction of movement that conscious sensory feedback and
adjustment of motor activity are virtually precluded, at least
until the act is nearly completed. Learning an element of a skill
thus involves developing a computer-program-like schedule of neural
activations that can be called up, so to speak, to effect a particular
desired end product of motor activity. Of course, the raw program
for a particular voluntary action is not sufficient to permit
the execution of that action: information regarding such parameters
as intended magnitude and direction of movement must be furnished
from the conscious sphere, and data indicating the position and
motion of the body platform relative to the surface of the earth--that
is, spatial orientation information--must be furnished from the
preconscious sphere. The necessity for the additional information
can be seen in the signature-writing example cited above: one
can write large or small, quickly or slowly, and on a horizontal
or vertical surface. Obviously, different patterns of neuromuscular
activation, even grossly different muscle groups, are needed to
accomplish a basic act under varying spatial and temporal conditions;
the necessary adjustments are made automatically, however, without
conscious intervention. Vestibular and other sensory data providing
spatial orientation information for use in either skilled voluntary
or reflexive motor activity are processed into a
preconscious
orientational percept that provides the informational basis upon
which such automatic adjustments are made. Thus, one can decide
what the outcome of his or her action is to be and initiate the
command to do it, without consciously having to discern the direction
of the force of gravity, analyze its potential effects on planned
motor activity, select appropriate muscle groups and modes of
activation to compensate for gravity, and then activate and deactivate
each muscle in proper sequence and with proper timing to accomplish
the desired motor activity. The body takes care of the details,
using stored programs for elements of skilled motor activity,
and the current preconscious orientational percept. This whole
process is the major function and responsibility of the vestibulocerebellar
axis.
Conscious Percepts
Usually as a result of the same
information processing that provides the preconscious orientational
percept, one also is provided a conscious orientational percept.
This percept can be false, that is, illusory, in which case the
individual is said to experience an orientational illusion, or
to have spatial disorientation. We can be aware, moreover, that
what our bodies tell us about our spatial orientation is not what
can be concluded from other information such as flight instrument
data. Conscious orientational percepts thus can be either natural
or derived, depending on the source of the orientation information
and the perceptual process involved; and an individual can experience
both natural and derived conscious orientational percepts at the
same time. Consequently, pilots who have become disoriented in
flight commonly exhibit vacillating control inputs, as they alternate
indecisively between responding first to one percept and then
to the other.
Figure
12. Ocular countertorsion, a vestibulo-ocular reflex of otolith-organ
origin. When the head is tilted to the left, the eyes rotate
to the right to assume a new angular position about the visual
axes, as shown.
Thresholds of Vestibular Perception
Often an orientational illusion
occurs because the physical event resulting in a change in bodily
orientation is below the threshold of perception. For that reason,
the student of disorientation should be aware of the approximate
perceptual thresholds associated with the various modes of vestibular
stimulation.
The lowest reported threshold
for perception of rotation is 0.035°/sec2, but this
degree of sensitivity is obtained only with virtually continuous
angular acceleration and long response latencies (20 to 40 seconds). 13
Other observations put the perceptual threshold
between roughly 0.1 and 2.0°/sec2; reasonable values
are 0.14, 0.5, and 0.5°/sec2 for yaw, roll, and pitch
motions, respectively.14 It is common practice, however,
to describe the thresholds of the semicircular ducts in terms
of the angular acceleration-time product, or angular velocity,
which results in just perceptible rotation. This product, known
as Mulder's constant, remains fairly constant for stimulus times
of about 5 seconds or less. Using the reasonable value of 2°/sec
for Mulder's constant, an angular acceleration of 50/sec2 applied for half
a second would be perceived because the acceleration-time
product is above the 2°/sec angular velocity threshold. But a
10°/sec2 acceleration applied for a tenth of a second
would not be perceived because it would be below the angular velocity
threshold; nor would a 0.2°/sec2 acceleration applied
for 5 seconds be perceived. Inflight experiments have shown that
blindfolded pilot subjects are not able to consistently perceive
roll rates of 1.0°/sec or less, but can perceive a roll when the
velocity is 2.0°/sec or higher. Pitch rate thresholds in flight
are also between 1.0 and 2.0°/sec. But when aircraft pitch motions
are coupled with compensatory power adjustments to keep the net
G force always directed toward the aircraft floor, the pitch threshold
is raised well above 2.0°/sec.15
The perceptual threshold related
to otolith-organ function necessarily involves both an angle and
a magnitude because the otolith organs respond to linear accelerations
and gravitoinertial forces, both of which have direction and intensity.
A 1.5° change in direction of applied G force is perceptible under
ideal (experimental) conditions. The minimum perceptible intensity
of linear acceleration has been reported by various authors to
be between 0.001 and 0.03 g, depending on the direction of acceleration
and the experimental method used. Values of 0.01 g for az and 0.006 g for
ax accelerations are appropriate representative
thresholds, and a similar value for ay acceleration is probably
reasonable. Again, these absolute thresholds apply when the acceleration
is either sustained or applied at relatively low frequencies.
The threshold for linear accelerations applied for less than about
5 seconds is a more or less constant acceleration-time product,
or linear velocity, of about 0.3 to 0.4 m/sec.
Unfortunately for those who would
like to calculate exactly what orientational percept results from
a particular set of linear and angular accelerations (e.g., those
occurring prior to an aircraft mishap), the actual vestibular
perceptual thresholds are, as expressed by one philosopher, "constant
except when they vary." Probably the most common reason for an
orientational perceptual threshold to be raised is inattention
to orientational cues because attention is directed to something
else. Other reasons might be a low state of mental arousal, fatigue,
drug effects, or innate individual variation. Whatever the reason,
it appears that individuals can monitor their orientation with
considerable sensitivity under some circumstances and with relative
insensitivity under others, which inconsistency can itself lead
to perceptual errors that result in orientational illusions.
Of paramount importance in the
generation of orientational illusions, however, is not the fact
that absolute vestibular thresholds exist or that vestibular thresholds
are time-varying. Rather, it is the fact that the components of
the vestibular system, like any complex mechanical or electrical
system, have characteristic frequency responses; and stimulation
by patterns of acceleration outside the optimal, or "design,"
frequency-response ranges of the semicircular ducts and otolith
organs causes the vestibular system to make errors. In night,
much of the stimulation resulting from the acceleratory environment
is indeed outside the design frequency-response ranges of the
vestibular end-organs; consequently, orientational illusions occur
in flight. Elucidation of this important point is provided in
the section entitled "Spatial Disorientation."
Vestibular Suppression and
Enhancement
Like all sensory systems, the
vestibular system exhibits a decreased response to stimuli that
are persistent (adaptation) or repetitious (habituation). Even
more important to the aviator is the fact that, with time and
practice, one can develop the ability to suppress natural vestibular
responses, both perceptual and motor. This ability is termed vestibular
suppression.16 Closely related to the concept of vestibular
suppression is that of visual dominance, the ability to obtain
and use spatial orientation cues from the visual environment despite
the presence of potentially strong vestibular cues. Vestibular
suppression seems to be exerted, in fact, through visual dominance
because it disappears in the absence of vision.17 The
opposite effect, that of an increase in perceptual and motor responsiveness
to vestibular stimulation, is termed vestibular enhancement. Such
enhancement can occur (1) when the stimulation is novel, as in
an amusement park ride; (2) threatening, as in an aircraft spinning
out of control; or (3) whenever spatial orientation is perceived
to be especially important. It is tempting to attribute to the
efferent vestibular neurons the function of controlling the gain
of the vestibular system so as to effect suppression and enhancement,
and some evidence exists to support that notion.18 The actual mechanisms
involved appear to be much more complex
than would be necessary merely to provide gross changes in the
gain of the vestibular end-organs. Precise control of vestibular
responses to anticipated stimulation, based on sensory efferent
copies of voluntary commands for movement, is probably exercised
by the cerebellum via a feed-forward loop involving the vestibular
efferent system. Thus, when discrepancies between anticipated
and actual stimulation generate a neural error signal, a response
is evoked, and vestibular reflexes and heightened perception occur.19
Vestibular suppression, then, involves the development of accurate
estimates of vestibular responses to orientational stimuli repeatedly
experienced, and the active countering of anticipated responses
by spatially and temporally patterned sensory efferent activity.
Vestibular enhancement, on the other hand, results from the lack
of available estimates of vestibular responses because of the
novelty of the stimulation, or perhaps from a revision in neural
processing strategy obligated by the failure of normal negative
feed-forward mechanisms to provide adequate orientation information.
Such marvelous complexity of vestibular function assures adaptability
to a wide variety of motional environments and thereby promotes
survival in them.
Thus, the sensory input from the
muscle spindles can be biased by descending influences from higher
neural centers such as the vestibulocerebellar axis. Thus, the
sensory input from the muscle spindles can be biased by descending
influences from higher neural centers such as the vestibulocerebellar
axis.
Although the muscle spindles are
structurally and functionally in parallel with associated muscle
groups and respond to changes in their length, the Golgi tendon
organs (Fig. 13b) are functionally in series with the muscles
and respond to changes in tension. A tendon organ consists of
a fusiform bundle of small tendon fascicles with intertwining
neural elements, and is located at the musculotendinous junction
or wholly within the tendon. Unlike that of the muscle spindle,
its innervation is entirely afferent.
The major function of both the
muscle spindles and the tendon organs is to provide the sensory
basis for myotatic (or muscle stretch) reflexes. These elementary
spinal reflexes operate to stabilize a joint by providing, in
response to an increase in length of a muscle and concomitant
stimulation of its included spindles, monosynaptic excitation
and contraction of the stretched agonist (e.g., extensor) muscle
and disynaptic inhibition and relaxation of its antagonist (e.g.,
flexor) muscle through the action of an inhibitory interneuron.
In addition, tension developed on associated tendon organs results
in disynaptic inhibition of the agonist muscle, thus regulating
the amount of contraction generated. The myotatic reflex mechanism
is, in fact, the foundation of posture and locomotion. Modification
of this and other basic spinal reflexes by organized facilitatory
or inhibitory intervention originating at higher neural levels,
either through direct action on skeletomotor (alpha) neurons or
through stimulation of fusimotor (primarily gamma) neurons to
muscle spindles, results in sustained postural equilibrium and
other purposive motor behavior. Some researchers have speculated,
moreover, that in certain types of spatial disorientation in flight,
this organized modification of spinal reflexes is interrupted
as cerebral cortical control of motor activity is replaced by
lower brainstem and spinal control. Perhaps the "frozen-on-the-controls"
type of disorientation-induced deterioration of flying ability
is a reflection of primitive reflexes made manifest by disorganization
of higher neural functions.
Despite the obvious importance
of the muscle spindles and tendon organs in the control of motor
activity, there is little evidence to indicate that their responding
to orientational stimuli (such as occur when one stands vertically
in a I-G environment) results in any corresponding conscious proprioceptive
percept.22 Nevertheless, it is known that the dorsal
columns and other ascending spinal tracts carry muscle afferent
information to medullary and thalamic relay nuclei and thence
to the cerebral sensory cortex. Furthermore, extensive projections
into the cerebellum, via dorsal and ventral spinocerebellar tracts,
ensure that proprioceptive information from the afferent terminations
of the muscle spindles and tendon organs is integrated with other
orientational information and is relayed to the vestibular nuclei,
cerebral cortex, and elsewhere as needed.
Joint Sensation
In contrast to the situation with
the so-called "muscle sense of position" just discussed, it has
been well established that sensory information from the joints
does reach consciousness. In fact, the threshold for perception
of joint motion and position can be quite low, as low as 0.5°
for the knee joint when moved at greater than 1.0°/sec. The receptors
in the joints are of three types, as shown in Figure 13c: (1)
lamellated or encapsulated Pacinian corpuscle-like end-organs;
(2) spray-type structures, known as Ruffini-like endings when
found in joint capsules and Golgi tendon organs when found in
ligaments; and (3) free nerve endings. The Pacinian corpuscle-like
terminals are rapidly adapting and are sensitive to quick movement
of the joint, whereas both of the spray-type endings are slowly
adapting and serve to signal slow joint movement and joint position.
There is evidence that polysynaptic spinal reflexes can be elicited
by stimulation of joint receptors, but their nature and extent
are not well understood. Proprioceptive information from the joint
receptors projects via the dorsal funiculi eventually to the cerebral
sensory cortex and via the spinocerebellar tracts to the anterior
lobe of the cerebellum.
One must not infer from this discussion
that only muscles, tendons, and joints have proprioceptive sensory
receptors. Both lamellated and spray-type receptors, as well as
free nerve endings, are found in fascia, aponeuroses, and other
connective tissues of the musculoskeletal system, and they presumably
provide proprioceptive information to the central nervous system
as well.
Cutaneous Exteroceptors
The exteroceptors of the skin
include: (1) mechanoreceptors, which respond to touch and pressure;
(2) thermoreceptors, which respond to heat and cold; and (3) nociceptors,
which respond to noxious mechanical and/or thermal events and
give rise to sensations of pain. Of the cutaneous exteroceptors,
only the mechanoreceptors contribute significantly to spatial
orientation.
A variety of receptors are involved
in cutaneous mechanoreception: spray-type Ruffini corpuscles,
lamellated Pacinian and Meissner corpuscles, branched and straight
lanceolate terminals, Merkel cells, and free nerve endings (Fig.
13d). The response patterns of mechanoreceptors also are numerous:
eleven different types of response, varying from high-frequency
transient detection through several modes of velocity detection
to more or less static displacement detection, have been recognized.
Pacinian corpuscles and certain receptors associated with
hair
follicles are very rapidly adapting and have the highest mechanical
frequency responses, responding to sinusoidal skin displacements
in the range of 50 to 400 Hz. They are thus well suited to monitor
vibration and transient touch stimuli. Ruffini corpuscles are
slowly adapting and, therefore, respond primarily to sustained
touch and pressure stimuli. Merkel cells appear to have a moderately
slowly adapting response, making them suitable for monitoring
static skin displacement and velocity. Meissner corpuscles seem
to detect primarily velocity of skin deformation. Other receptors
provide other types of response, so as to complete the spectrum
of mechanical stimuli that can be sensed through the skin. The
mechanical threshold for the touch receptors is quite low--less
than 0.03 dyne/cm2 on the thumb. (In comparison with the labyrinthine
receptors subserving audition, however, this threshold is not
so impressive: a O-dB sound pressure level represents 0.0002 dyne/cm2,
more than 100 times lower.) Afferent information from the described
mechanoreceptors is conveyed to the cerebral cortex mainly by
way of the dorsal funiculi and medullary relay nuclei into the
medial lemnisci and thalamocortical projections. The dorsal spinocerebellar
tract and other tracts to the cerebellum provide the pathways
by which cutaneous exteroceptive information reaches the cerebellum
and is integrated with proprioceptive information from muscles,
tendons, joints, and vestibular end-organs.
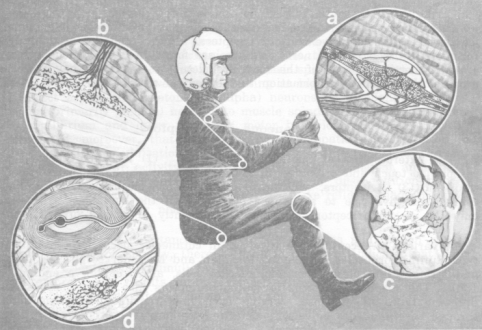
Figure
13. Some of the nonvestibular proprioceptive and cutaneous exteroceptive
receptors sub serving spatial orientation. a. Muscle spindle,
with central afferent (sensory) and more peripheral efferent (fusimotor)
innervations. b. Golgi tendon organ. c. Lamellated, spray-type,
and free-nerve-ending joint receptors. d. Two of the many types
of mechanoreceptors found in the skin: lamellated Pacinian corpuscles
and spray-type Ruffini corpuscles.
AUDITORY ORIENTATION
On the surface of the earth, the
ability to determine the location of a sound source can play a
role in spatial orientation, as evidenced by the fact that a revolving
sound source can create a sense of self-rotation and even elicit
reflex compensatory and anticompensatory eye movements called
audiokinetic nystagmus. Differential filtering of incident sound
energy by the external ear, head, and shoulders at different relative
locations of the sound source provides the ability to discriminate
sound location. Part of this discrimination process involves analysis
of interaural differences in arrival time of congruent sounds;
but direction-dependent changes in spectral characteristics of
incident sound energies allow the listener to localize sounds
in elevation and azimuth (and to some extent range), even when
the interaural arrival times are not different. In
aircraft, binaural
sound localization is of little use in spatial orientation because
of high ambient noise levels and the absence of audible external
sound sources. Pilots do extract some orientation information,
however, from the auditory cues provided by the rush of air past
the airframe: the sound frequencies and intensities characteristic
of various airspeeds and angles of attack are recognized by the
experienced pilot, who uses them in conjunction with other orientation
information to create a percept of velocity and pitch attitude
of the aircraft. As aircraft have become more capable, however,
and the pilot has become more insulated from such acoustic stimuli,
the usefulness of aircraft-generated auditory orientation cues
has diminished.